
Exercising D. melanogaster Modulates the Mitochondrial Proteome and Physiology. The Effect on Lifespan Depends upon Age and Sex

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fly Husbandry
2.2. Fly Separation
2.3. Exercise Regime
2.4. Mortality
2.5. Climbing Assay
2.6. Tissue Preparation for HRR
2.7. High-Resolution Respirometry
2.8. Mitochondrial Preparation
2.9. 2D Gel Electrophoresis (2D–PAGE)
2.10. Matrix-Assisted Laser Desorption Ionization Tandem Time-of-Flight Mass Spectrometry (MALDI–TOF/MS)
2.11. Liquid Chromatography Tandem Mass Spectrometry (LC–MS/MS)
2.12. Bioinformatic Analyses
3. Results
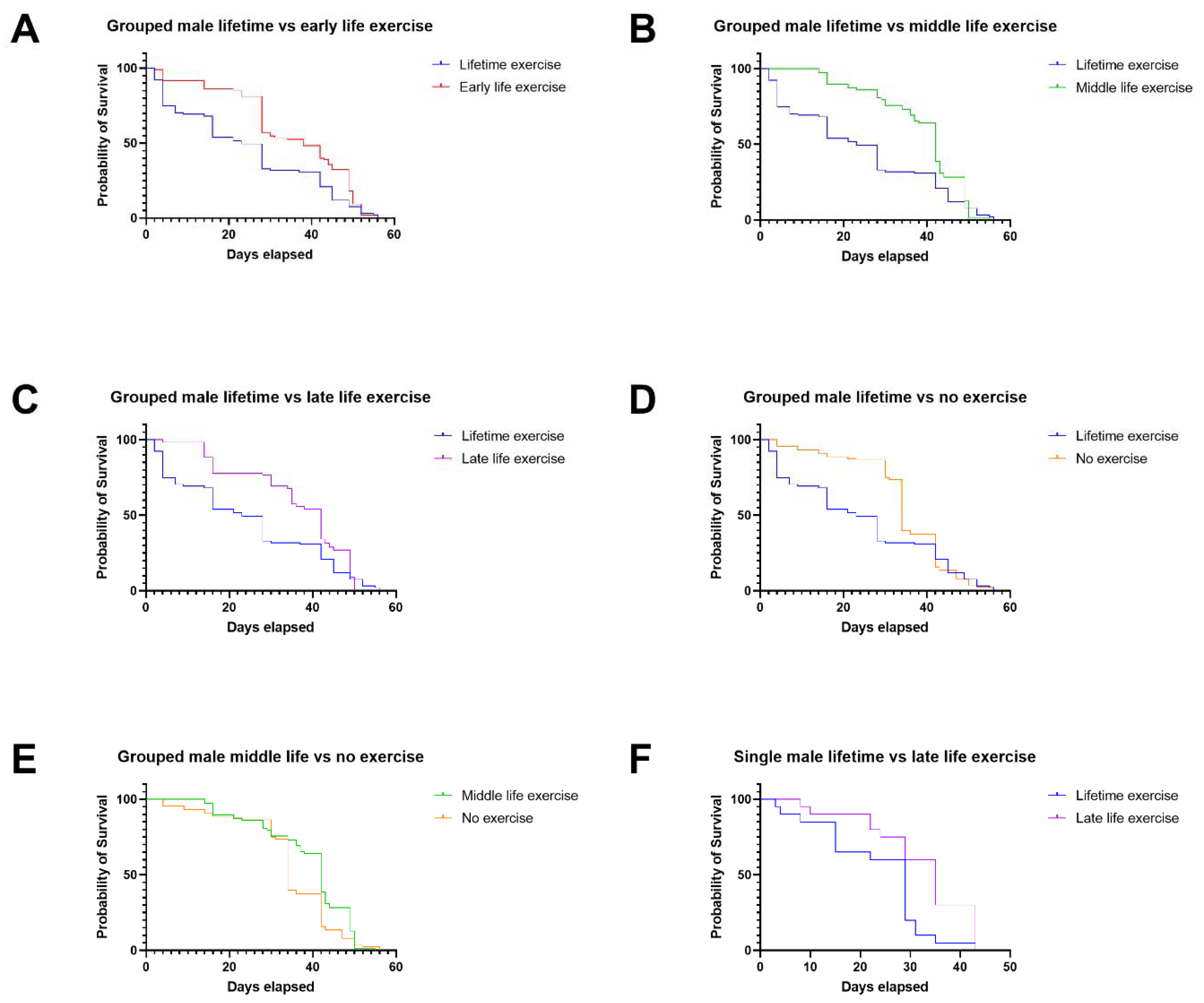
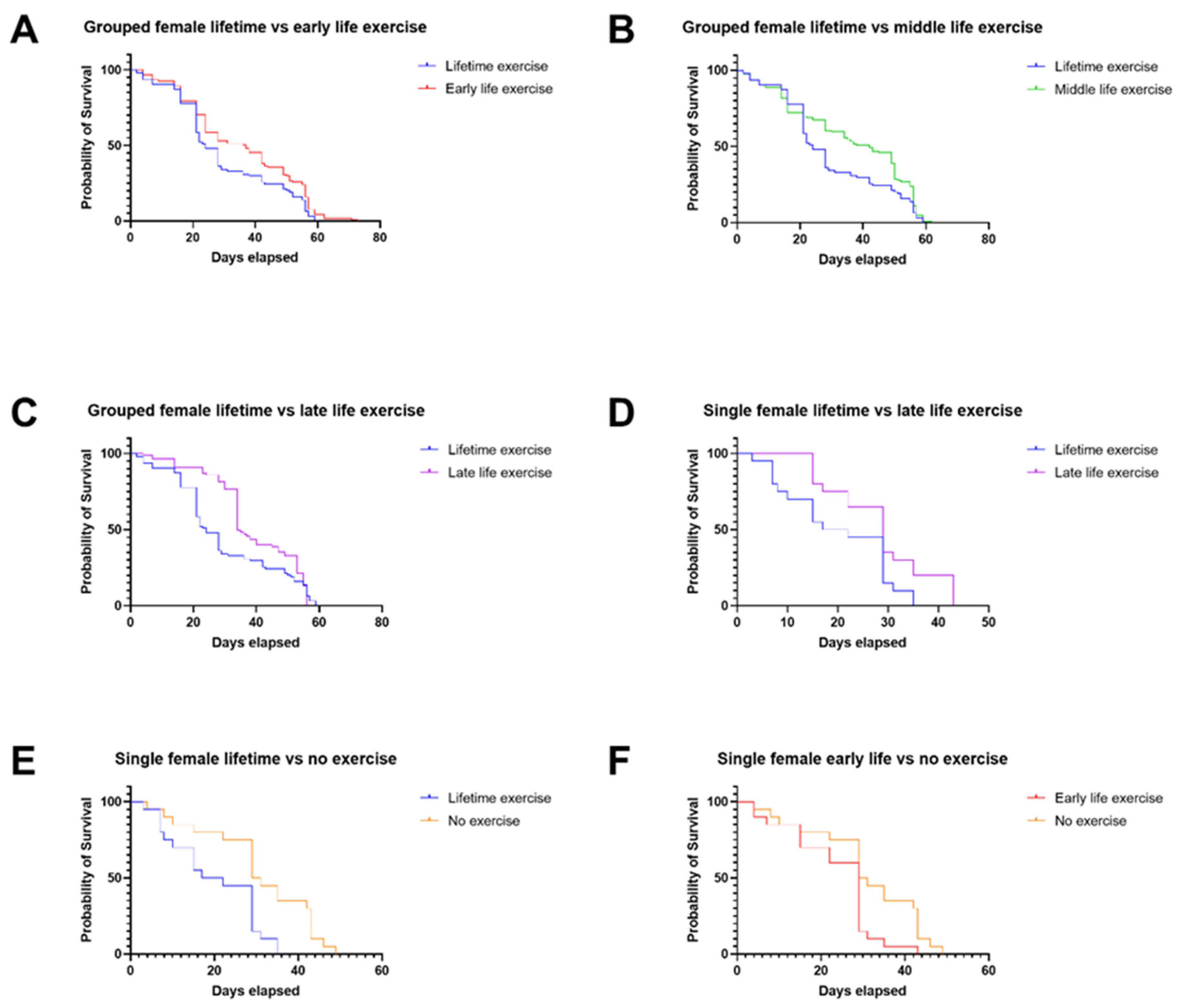
3.1. In Males, Late-Life Exercise Has the Most Beneficial Effect, and Exercise throughout Life Is Detrimental in Comparison
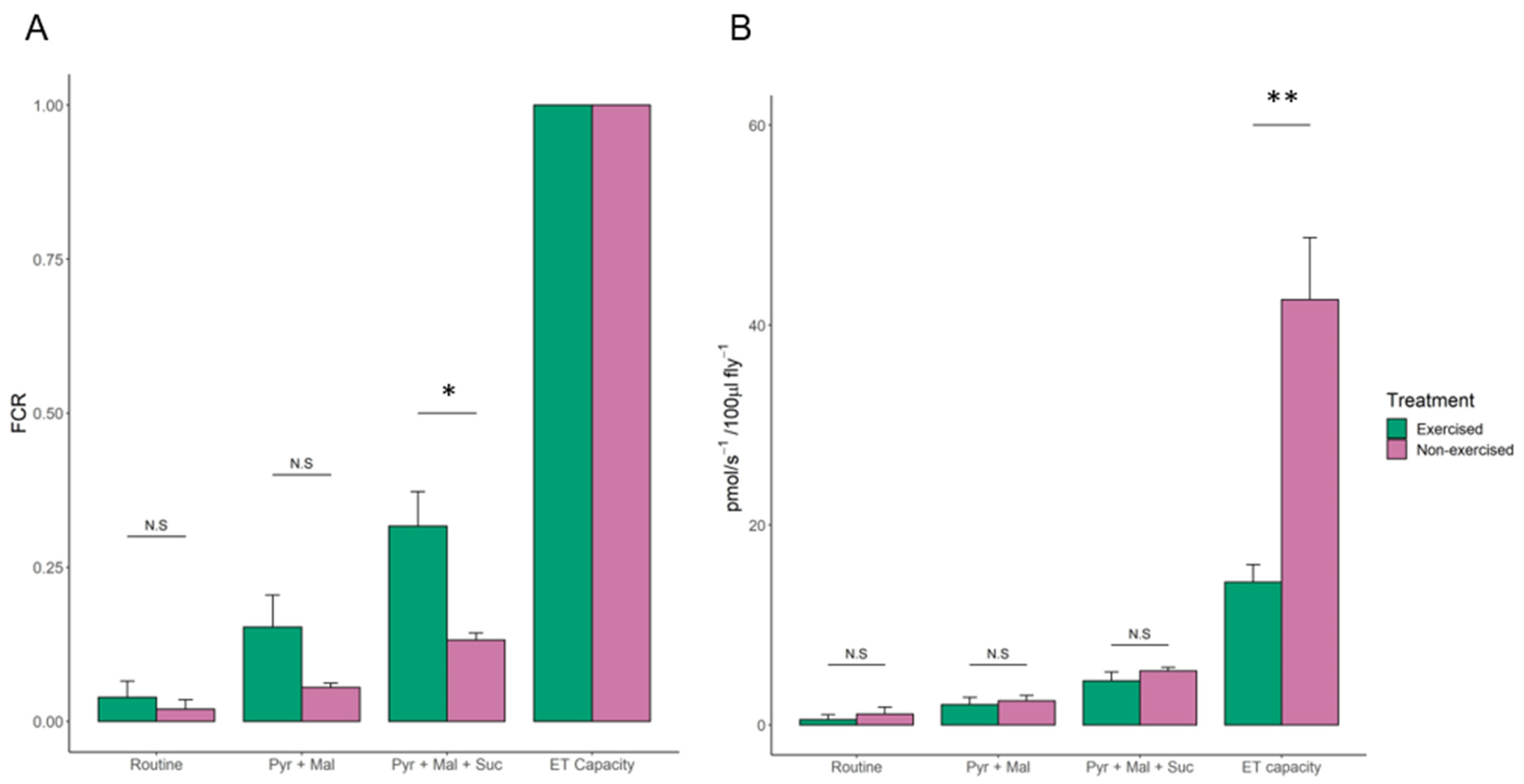
3.2. High-Resolution Respirometry of Exercised Drosophila
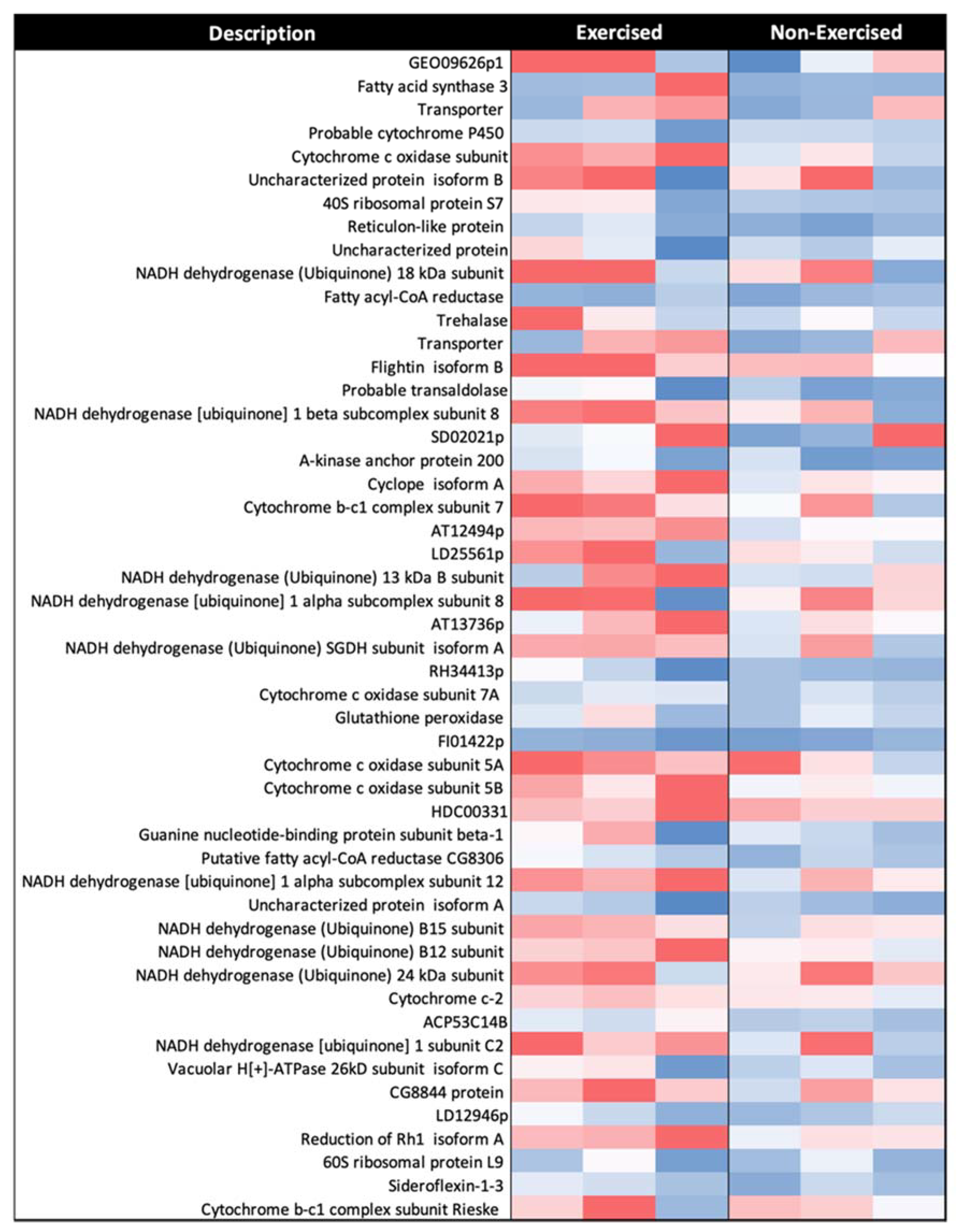
3.3. There Is Increased Expression of Many Proteins in Response to Exercise, including Those of the Electron Transport Chain
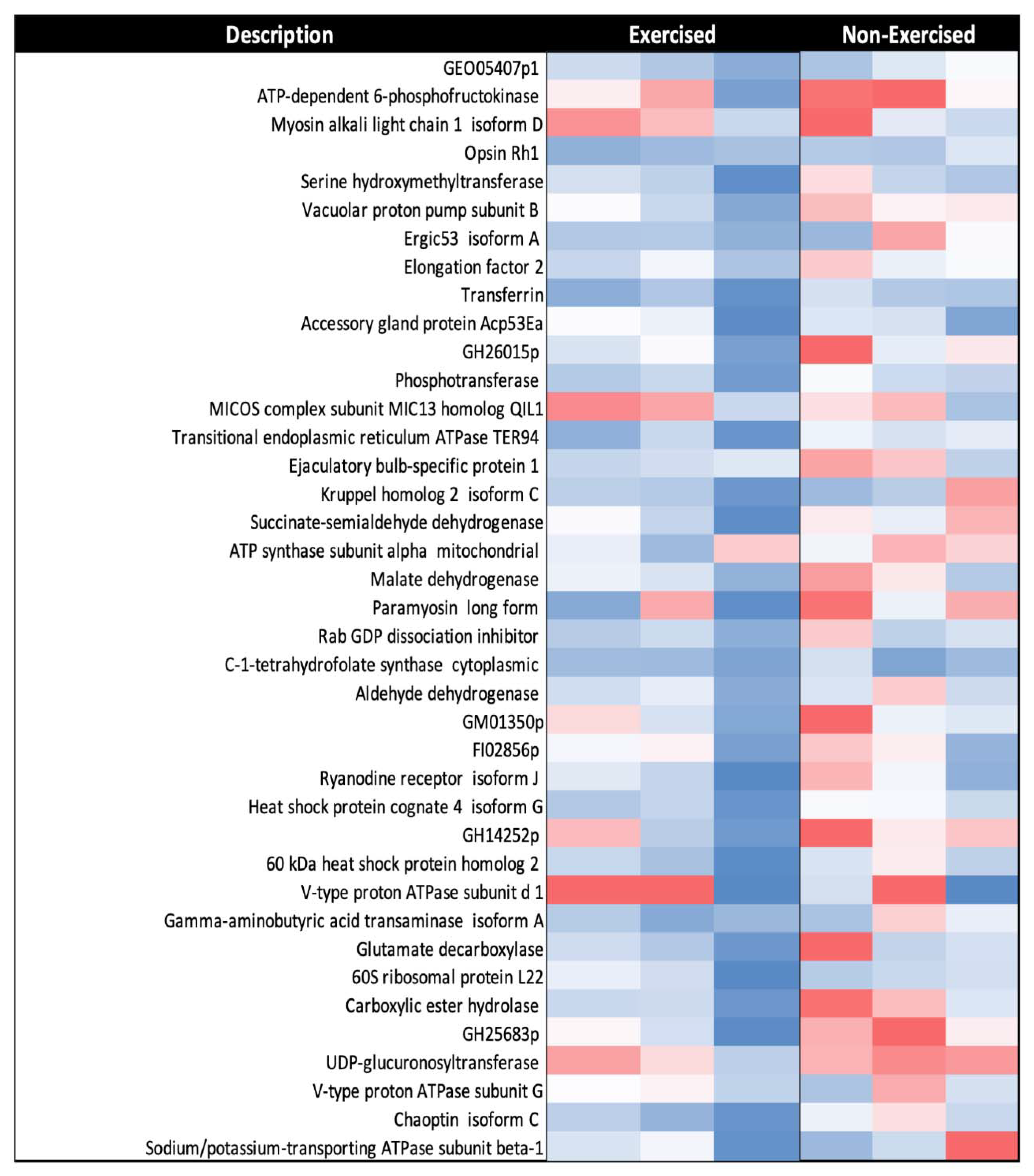
3.4. A Subset of Mitochondrial Proteins Decreased in Response to One Week of Exercise in D. melanogaster
4. Discussion
4.1. Lifetime Exercise Has Worse Outcomes Than Targeted Exercise Interventions in Male and Female Drosophila
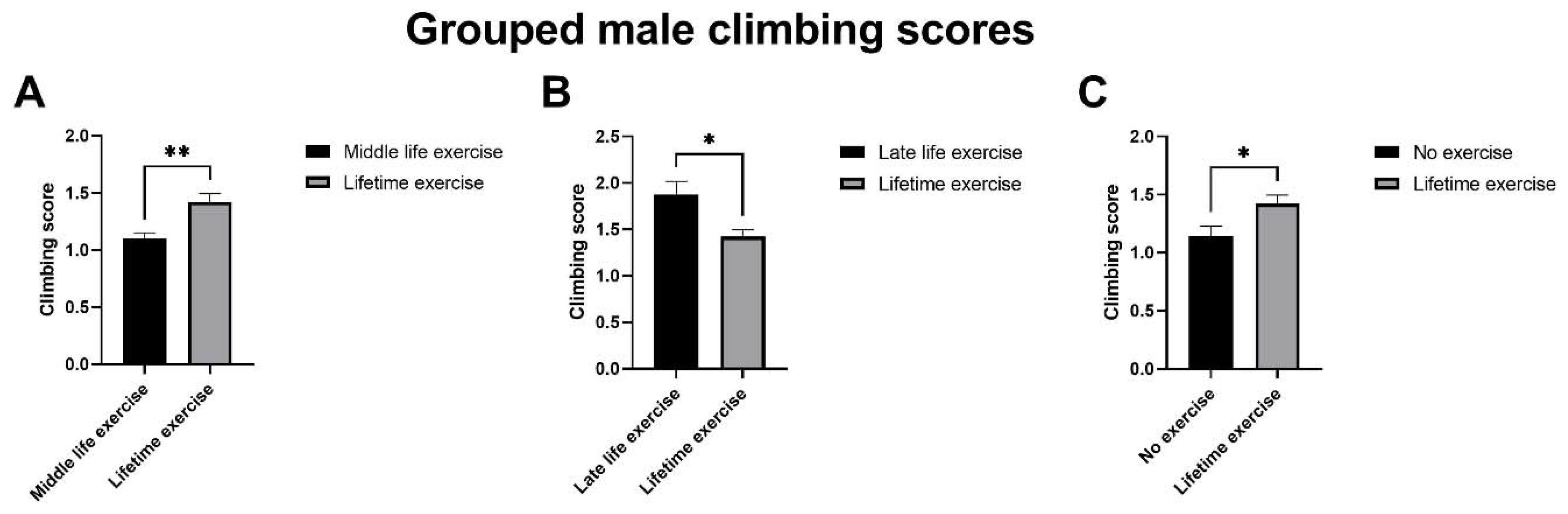
4.2. Late-Life Exercise Produces a Rapid Improvement in Climbing Assay Scores Compared with Life-Time Exercise for Grouped Male Drosophila
4.3. Succinate-Linked Respiration Is Elevated in Exercised Flies
4.4. Reduced Spare Respiratory Capacity in Exercised Drosophila May Promote Longevity
4.5. Proteins from the Electron Transport Chain Are Significantly Upregulated in Response to Exercise
4.6. Multiple Metabolic Pathways Are Downregulated in Response to Exercise
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mitchell, E.; Walker, R. Global ageing: Successes, challenges and opportunities. Br. J. Hosp. Med. 2020, 81, 1–9. [Google Scholar] [CrossRef]
- Marešová, P.; Mohelská, H.; Kuča, K. Economics Aspects of Ageing Population. Procedia Econ. Financ. 2015, 23, 534–538. [Google Scholar] [CrossRef] [Green Version]
- Kaeberlein, M. How healthy is the healthspan concept? GeroScience 2018, 40, 361–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austad, S.N. Sex Differences in Longevity and Aging. In Handbook of the Biology of Aging; Elsevier: Amsterdam, The Netherlands, 2011; pp. 479–495. [Google Scholar]
- Brent, L.J.N.; Ruiz-Lambides, A.; Platt, M.L. Family network size and survival across the lifespan of female macaques. Proc. R. Soc. B Boil. Sci. 2017, 284, 20170515. [Google Scholar] [CrossRef]
- Stanton, M.A.; Mann, J. Early Social Networks Predict Survival in Wild Bottlenose Dolphins. PLoS ONE 2012, 7, e47508. [Google Scholar] [CrossRef]
- Silk, J.B.; Beehner, J.; Bergman, T.J.; Crockford, C.; Engh, A.L.; Moscovice, L.R.; Wittig, R.M.; Seyfarth, R.M.; Cheney, D.L. Strong and Consistent Social Bonds Enhance the Longevity of Female Baboons. Curr. Biol. 2010, 20, 1359–1361. [Google Scholar] [CrossRef] [Green Version]
- Holt-Lunstad, J.; Smith, T.B.; Layton, J.B. Social Relationships and Mortality Risk: A Meta-analytic Review. PLoS Med. 2010, 7, e1000316. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, L.; Richardson, D.S.; Eikenaar, C.; Komdeur, J. The role of group size and environmental factors on survival in a cooperatively breeding tropical passerine. J. Anim. Ecol. 2006, 75, 1321–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gager, Y.; Gimenez, O.; O’Mara, M.T.; Dechmann, D.K.N. Group size, survival and surprisingly short lifespan in socially foraging bats. BMC Ecol. 2016, 16, 2. [Google Scholar] [CrossRef] [Green Version]
- Blumstein, D.T.; Williams, D.M.; Lim, A.N.; Kroeger, S.; Martin, J. Strong social relationships are associated with decreased longevity in a facultatively social mammal. Proc. R. Soc. B Boil. Sci. 2018, 285, 20171934. [Google Scholar] [CrossRef] [Green Version]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [Green Version]
- Spinelli, J.B.; Haigis, M.C. The multifaceted contributions of mitochondria to cellular metabolism. Nat. Cell Biol. 2018, 20, 745–754. [Google Scholar] [CrossRef]
- Giorgi, C.; Marchi, S.; Pinton, P. The machineries, regulation and cellular functions of mitochondrial calcium. Nat. Rev. Mol. Cell Biol. 2018, 19, 713–730. [Google Scholar] [CrossRef]
- Ward, D.M.; Cloonan, S.M. Mitochondrial Iron in Human Health and Disease. Annu. Rev. Physiol. 2019, 81, 453–482. [Google Scholar] [CrossRef]
- Martínez-Reyes, I.; Chandel, N.S. Mitochondrial TCA cycle metabolites control physiology and disease. Nat. Commun. 2020, 11, 102. [Google Scholar] [CrossRef] [Green Version]
- Letts, J.; Fiedorczuk, K.; Sazanov, L. The architecture of respiratory supercomplexes. Nat. Cell Biol. 2016, 537, 644–648. [Google Scholar] [CrossRef]
- Wang, C.; Youle, R.J. The Role of Mitochondria in Apoptosis. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Ebanks, B.; Ingram, T.L.; Chakrabarti, L. ATP synthase and Alzheimer’s disease: Putting a spin on the mitochondrial hypothesis. Aging 2020, 12, 16647–16662. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Halliwell, B. Oxidative stress, dysfunctional glucose metabolism and Alzheimer disease. Nat. Rev. Neurosci. 2019, 20, 148–160. [Google Scholar] [CrossRef]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Primers 2017, 3, 1–21. [Google Scholar] [CrossRef]
- Chen, X.; Yi, Z.; Wong, G.T.; Hasan, K.M.M.; Kwan, J.S.; Ma, A.C.; Chang, R.C. Is exercise a senolytic medicine? A systematic review. Aging Cell 2021, 20, e13294. [Google Scholar] [CrossRef]
- Ristow, M.; Zarse, K.; Oberbach, A.; Klöting, N.; Birringer, M.; Kiehntopf, M.; Stumvoll, M.; Kahn, C.R.; Blüher, M. Antioxidants prevent health-promoting effects of physical exercise in humans. Proc. Natl. Acad. Sci. USA 2009, 106, 8665–8670. [Google Scholar] [CrossRef] [Green Version]
- Bárcena, C.; Mayoral, P.; Quirós, P.M. Mitohormesis, an Antiageing Paradigm. Int. Rev. Cell Mol. Biol. 2018, 340, 35–77. [Google Scholar]
- Sun, Y.; Yolitz, J.; Wang, C.; Spangler, E.; Zhan, M.; Zou, S. Aging Studies in Drosophila melanogaster. Methods Mol. Biol. 2013, 1048, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Moreno, M.A.; Farr, C.L.; Kaguni, L.S.; Garesse, R. Drosophila melanogaster as a Model System to Study Mitochondrial Biology. Methods Mol. Biol. 2007, 372, 33–49. [Google Scholar] [CrossRef] [Green Version]
- Plyusnina, E.N.; Shaposhnikov, M.V.; Moskalev, A.A. Increase of Drosophila melanogaster lifespan due to D-GADD45 overexpression in the nervous system. Biogerontology 2010, 12, 211–226. [Google Scholar] [CrossRef]
- Sun, J.; Molitor, J.; Tower, J. Effects of simultaneous over-expression of Cu/ZnSOD and MnSOD on Drosophila melanogaster life span. Mech. Ageing Dev. 2004, 125, 341–349. [Google Scholar] [CrossRef]
- Yang, Y.; Gehrke, S.; Imai, Y.; Huang, Z.; Ouyang, Y.; Wang, J.; Yang, L.; Beal, M.F.; Vogel, O.H.; Lu, B. Mitochondrial pathology and muscle and dopaminergic neuron degeneration caused by inactivation of Drosophila Pink1 is rescued by Parkin. Proc. Natl. Acad. Sci. USA 2006, 103, 10793–10798. [Google Scholar] [CrossRef] [Green Version]
- Clark, I.E.; Dodson, M.W.; Jiang, C.; Cao, J.H.; Huh, J.R.; Seol, J.H.; Yoo, S.J.; Hay, B.A.; Guo, M. Drosophila pink1 is required for mitochondrial function and interacts genetically with parkin. Nat. Cell Biol. 2006, 441, 1162–1166. [Google Scholar] [CrossRef]
- Park, J.; Lee, S.B.; Lee, S.; Kim, Y.; Song, S.; Kim, S.; Bae, E.; Kim, J.; Shong, M.; Kim, J.-M.; et al. Mitochondrial dysfunction in Drosophila PINK1 mutants is complemented by parkin. Nat. Cell Biol. 2006, 441, 1157–1161. [Google Scholar] [CrossRef]
- Kasarda, D.D.; Tao, H.P.; Evans, P.K.; Adalsteins, A.E.; Yuen, S.W. Sequencing of Protein from a Single Spot of a 2-D Gel Pattern: N-Terminal Sequence of a Major Wheat LMW-Glutenin Subunit. J. Exp. Bot. 1988, 39, 899–906. [Google Scholar] [CrossRef]
- Zhu, W.; Smith, J.W.; Huang, C.-M. Mass Spectrometry-Based Label-Free Quantitative Proteomics. J. Biomed. Biotechnol. 2009, 2010, 840518. [Google Scholar] [CrossRef]
- Tinkerhess, M.J.; Healy, L.; Morgan, M.; Sujkowski, A.; Matthys, E.; Zheng, L.; Wessells, R. The Drosophila PGC-1α Homolog spargel Modulates the Physiological Effects of Endurance Exercise. PLoS ONE 2012, 7, e31633. [Google Scholar] [CrossRef]
- Gargano, J.W.; Martin, I.; Bhandari, P.; Grotewiel, M.S. Rapid iterative negative geotaxis (RING): A new method for assessing age-related locomotor decline in Drosophila. Exp. Gerontol. 2005, 40, 386–395. [Google Scholar] [CrossRef]
- Shephard, F.; Greville-Heygate, O.; Liddell, S.; Emes, R.; Chakrabarti, L. Analysis of Mitochondrial haemoglobin in Parkinson’s disease brain. Mitochondrion 2016, 29, 45–52. [Google Scholar] [CrossRef]
- Fisher-Wellman, K.; Bloomer, R.J. Acute exercise and oxidative stress: A 30 year history. Dyn. Med. 2009, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Hekimi, S.; Lapointe, J.; Wen, Y. Taking a “good” look at free radicals in the aging process. Trends Cell Biol. 2011, 21, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Golden, T.R.; Hinerfeld, D.A.; Melov, S. Oxidative stress and aging: Beyond correlation. Aging Cell 2002, 1, 117–123. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; DELLA Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Sujkowski, A.; Ramesh, D.; Brockmann, A.; Wessells, R. Octopamine Drives Endurance Exercise Adaptations in Drosophila. Cell Rep. 2017, 21, 1809–1823. [Google Scholar] [CrossRef] [Green Version]
- Gorbacheva, T.M.; Syromyatnicov, M.Y.; Popov, V.N.; Lopatin, A.V.; Eprintsev, A.T.; Fedorin, D.N. Characteristics of functioning of succinate dehydrogenase from flight muscles of the bumblebee Bombus terrestris (L.). Biol. Bull. 2013, 40, 429–434. [Google Scholar] [CrossRef]
- Zakharchenko, M.V.; Zakharchenko, A.; Khunderyakova, N.; Tutukina, M.; Simonova, M.; Vasilieva, A.; Romanova, O.; Fedotcheva, N.; Litvinova, E.; Maevsky, E.; et al. Burst of succinate dehydrogenase and α-ketoglutarate dehydrogenase activity in concert with the expression of genes coding for respiratory chain proteins underlies short-term beneficial physiological stress in mitochondria. Int. J. Biochem. Cell Biol. 2012, 45, 190–200. [Google Scholar] [CrossRef]
- Marchetti, P.; Fovez, Q.; Germain, N.; Khamari, R.; Kluza, J. Mitochondrial spare respiratory capacity: Mechanisms, regulation, and significance in non-transformed and cancer cells. FASEB J. 2020, 34, 13106–13124. [Google Scholar] [CrossRef]
- Silva, L.A.; Pinho, C.A.; Scarabelot, K.S.; Fraga, D.B.; Volpato, A.M.J.; Boeck, C.R.; De Souza, C.T.; Streck, E.L.; Pinho, R.A. Physical exercise increases mitochondrial function and reduces oxidative damage in skeletal muscle. Eur. J. Appl. Physiol. 2009, 105, 861–867. [Google Scholar] [CrossRef]
- Skinner, J.S.; McLellan, T.H. The Transition from Aerobic to Anaerobic Metabolism. Res. Q. Exerc. Sport 1980, 51, 234–248. [Google Scholar] [CrossRef]
- Tatarková, Z.; Kuka, S.; Račay, P.; Lehotsky, J.; Dobrota, D.; Mištuna, D.; Kaplan, P. Effects of Aging on Activities of Mitochondrial Electron Transport Chain Complexes and Oxidative Damage in Rat Heart. Physiol. Res. 2011, 60, 281–289. [Google Scholar] [CrossRef]
- Tavallaie, M.; Voshtani, R.; Deng, X.; Qiao, Y.; Jiang, F.; Collman, J.P.; Fu, L. Moderation of mitochondrial respiration mitigates metabolic syndrome of aging. Proc. Natl. Acad. Sci. USA 2020, 117, 9840–9850. [Google Scholar] [CrossRef]
- Sabath, N.; Levy-Adam, F.; Younis, A.; Rozales, K.; Meller, A.; Hadar, S.; Soueid-Baumgarten, S.; Shalgi, R. Cellular proteostasis decline in human senescence. Proc. Natl. Acad. Sci. USA 2020, 117, 31902–31913. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exercised | Non-Exercised | ||
|---|---|---|---|
| n = 5 | n = 4 | ||
| Stage | Mean (s.e) | Mean (s.e) | p-Value |
| Routine | |||
| Specific flux | 0.55 (0.48) | 1.08 (0.70) | 0.537 |
| FCR | 0.04 (0.03) | 0.02 (0.02) | 0.58 |
| Pyruvate & Malate | |||
| Specific flux | 2.04 (0.70) | 2.41 (0.54) | 0.699 |
| FCR | 0.15 (0.05) | 0.05 (0.01) | 0.143 |
| Succinate | |||
| Specific flux | 4.40 (0.89) | 5.39 (0.35) | 0.381 |
| FCR | 0.32 (0.06) | 0.13 (0.01) | 0.025 * |
| ET capacity | |||
| Specific flux | 14.27 (1.77) | 42.55 (6.19) | 0.002 ** |
| Description | Accession | Significance (−log10P) | RAR | Log2 FC | |
|---|---|---|---|---|---|
| Complex I | |||||
| NADH dehydrogenase (ubiquinone) | 1Subunit C2 | Q9VQM2 | 200 | 1.53 | 0.61 |
| 1 Beta subcomplex subunit 8 mitochondrial | Q9W3X7 | 133.11 | 1.85 | 0.89 | |
| 1 Alpha subcomplex subunit 8 | Q9W125 | 142.39 | 1.68 | 0.74 | |
| 1 Alpha subcomplex subunit 12 | Q9VQD7 | 200 | 1.57 | 0.65 | |
| SGDH subunit isoform A | Q9VTU2 | 86.17 | 1.64 | 0.71 | |
| B15 subunit | Q6IDF5 | 36.57 | 1.56 | 0.64 | |
| B12 subunit isoform A | Q9W2E8 | 88.95 | 1.55 | 0.64 | |
| 24 kDa subunit isoform A | Q9VX36 | 65.53 | 1.54 | 0.63 | |
| 18 kDa subunit | Q9VWI0 | 200 | 2.05 | 1.04 | |
| 13 kDa Bsubunit | Q9VTB4 | 78.35 | 1.69 | 0.76 | |
| GEO09626p1 | Q8SYJ2 | 26.03 | 2.92 | 1.55 | |
| EG:152A3.7 protein | O97418 | 29.75 | 1.9 | 0.93 | |
| CG8844 protein | Q9VQR2 | 200 | 1.53 | 0.61 | |
| AT12494p | Q9VJZ4 | 70.67 | 1.7 | 0.77 | |
| Complex III | |||||
| Cytochrome b-c1 complex | Subunit Rieske mitochondrial | Q9VQ29 | 80.35 | 1.5 | 0.59 |
| Subunit 7 | Q9VXI6 | 200 | 1.73 | 0.79 | |
| AT13736p | Q9VVH5 | 129.49 | 1.65 | 0.72 | |
| Complex IV | |||||
| Cyclope isoform A | Q9VMS1 | 200 | 1.78 | 0.83 | |
| Cytochrome C oxidase | Cytochrome C oxidase subunit | Q8IQW2 | 200 | 2.14 | 1.1 |
| Subunit 7A mitochondrial | Q9VHS2 | 82.59 | 1.61 | 0.69 | |
| Subunit 5A mitochondrial | Q94514 | 110.61 | 1.6 | 0.68 | |
| Subunit 5B isoform A | Q9VMB9 | 140.51 | 1.59 | 0.67 | |
| GEO09626p1 | Q8SYJ2 | 26.03 | 2.92 | 1.55 | |
| Other OXPHOS proteins | |||||
| HDC00331 | Q6IHY5 | 200 | 1.58 | 0.66 | |
| Uncharacterized protein isoform A | Q0KHZ6 | 93.18 | 1.56 | 0.64 | |
| Cytochrome c-2 | P84029 | 38.14 | 1.54 | 0.62 | |
| Description | Accession | Significance (−log10P) | RAR | Log2 FC |
|---|---|---|---|---|
| ATP metabolic process | ||||
| ATP synthase subunit alpha mitochondrial | P35381 | 84.91 | 0.62 | −0.68 |
| TCA cycle | ||||
| GM01350p | Q9VGQ1 | 32.66 | 0.58 | −0.78 |
| Malate dehydrogenase | Q9VKX2 | 56.18 | 0.62 | −0.68 |
| Pyruvate metabolic process | ||||
| Aldehyde dehydrogenase | Q9VLC5 | 38.53 | 0.59 | −0.76 |
| Cellular amino acid metabolic process | ||||
| Succinate-semialdehyde dehydrogenase | Q9VBP6 | 77.81 | 0.63 | −0.67 |
| Gamma-aminobutyric acid transaminase isoform A | Q9VW68 | 200 | 0.52 | −0.94 |
| Serine hydroxymethyltransferase | Q9W457 | 200 | 0.66 | −0.59 |
| Carboxylic acid metabolic process | ||||
| GM01350p | Q9VGQ1 | 32.66 | 0.58 | −0.78 |
| Aldehyde dehydrogenase | Q9VLC5 | 38.53 | 0.59 | −0.76 |
| Malate dehydrogenase | Q9VKX2 | 56.18 | 0.62 | −0.68 |
| Serine hydroxymethyltransferase | Q9W457 | 200 | 0.66 | −0.59 |
| Succinate-semialdehyde dehydrogenase | Q9VBP6 | 77.81 | 0.63 | −0.67 |
| Gamma-aminobutyric acid transaminase isoform A | Q9VW68 | 200 | 0.52 | −0.94 |
| Chaperone-mediated protein folding | ||||
| Heat shock protein cognate 4 isoform G | C7LA75 | 153.53 | 0.58 | −0.8 |
| 60 kDa heat shock protein homolog 2 mitochondrial | Q9VMN5 | 200 | 0.55 | −0.87 |
| Other mitochondrial proteins | ||||
| MICOS complex subunit MIC60 | A0A0B4KGN2 | 27.65 | 0.64 | −0.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ebanks, B.; Wang, Y.; Katyal, G.; Sargent, C.; Ingram, T.L.; Bowman, A.; Moisoi, N.; Chakrabarti, L. Exercising D. melanogaster Modulates the Mitochondrial Proteome and Physiology. The Effect on Lifespan Depends upon Age and Sex. Int. J. Mol. Sci. 2021, 22, 11606. https://doi.org/10.3390/ijms222111606
Ebanks B, Wang Y, Katyal G, Sargent C, Ingram TL, Bowman A, Moisoi N, Chakrabarti L. Exercising D. melanogaster Modulates the Mitochondrial Proteome and Physiology. The Effect on Lifespan Depends upon Age and Sex. International Journal of Molecular Sciences. 2021; 22(21):11606. https://doi.org/10.3390/ijms222111606
Chicago/Turabian StyleEbanks, Brad, Ying Wang, Gunjan Katyal, Chloe Sargent, Thomas L. Ingram, Antonia Bowman, Nicoleta Moisoi, and Lisa Chakrabarti. 2021. "Exercising D. melanogaster Modulates the Mitochondrial Proteome and Physiology. The Effect on Lifespan Depends upon Age and Sex" International Journal of Molecular Sciences 22, no. 21: 11606. https://doi.org/10.3390/ijms222111606