
Biological Control of Leaf Blight Disease Caused by Pestalotiopsis maculans and Growth Promotion of Quercus acutissima Carruth Container Seedlings Using Bacillus velezensis CE 100

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Antagonistic Activity of Bacillus velezensis CE 100 against Pestalotiopsis maculans
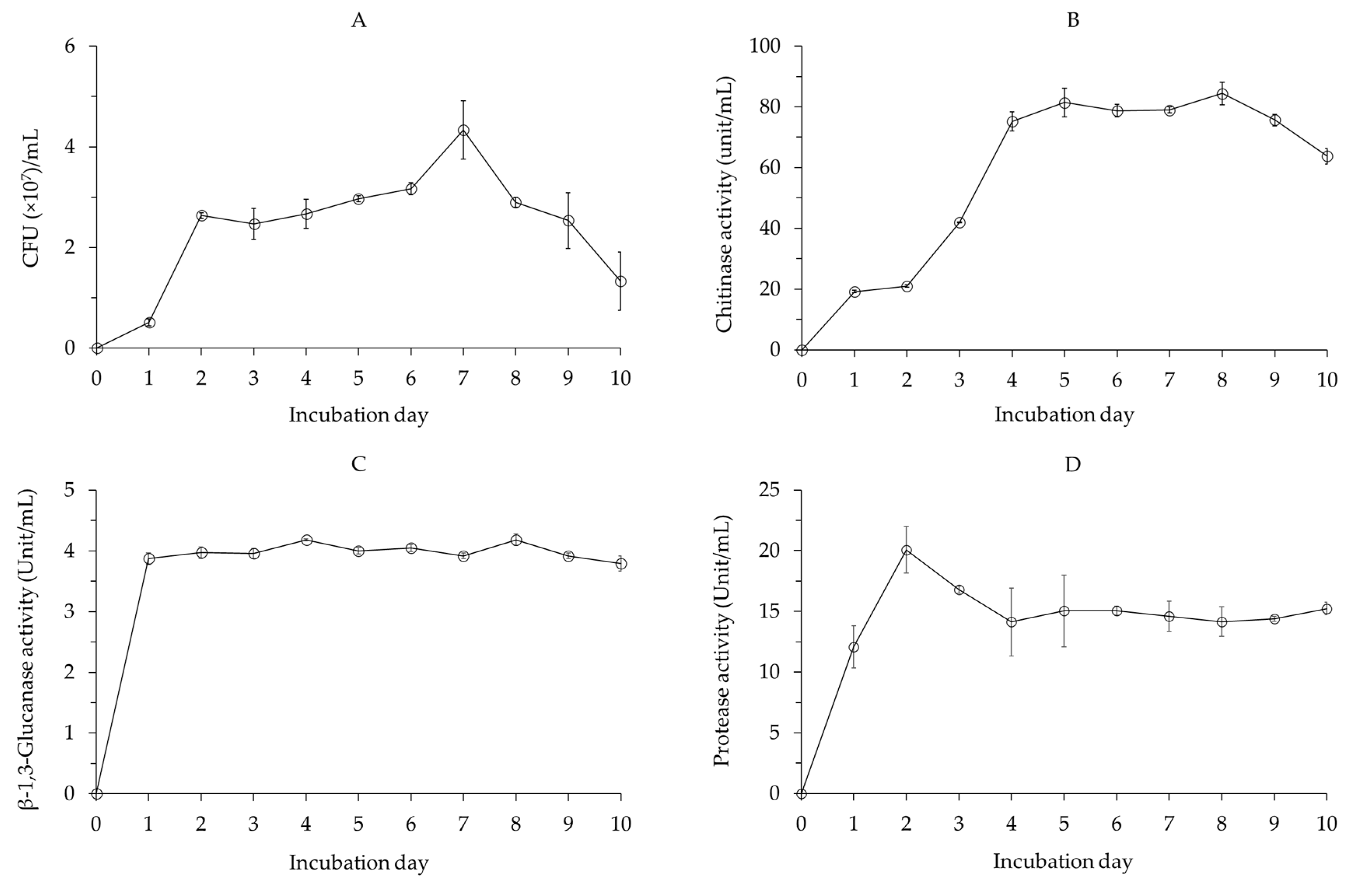
2.1.1. Cell Growth and Lytic Enzyme Production by Bacillus velezensis CE 100
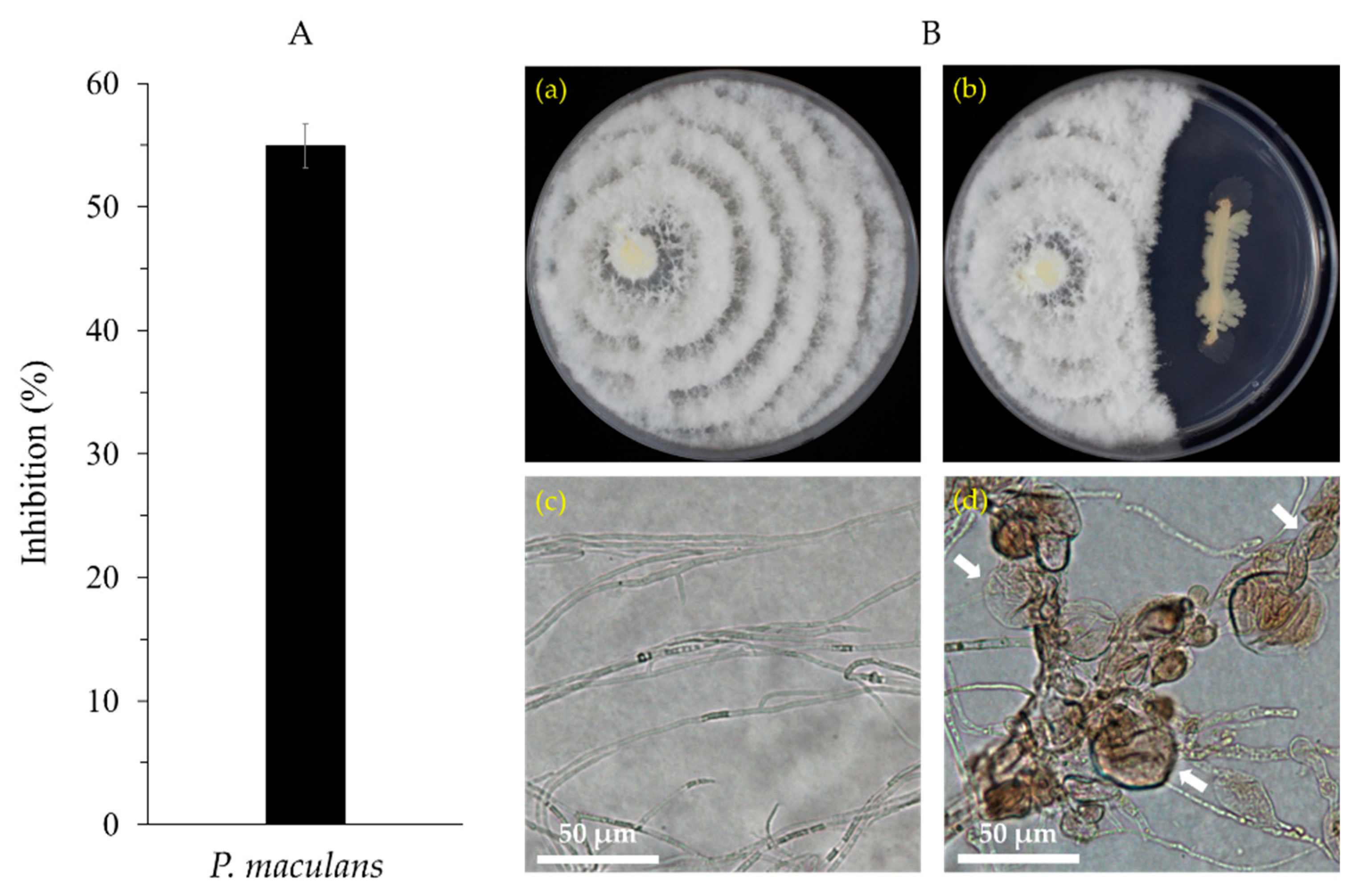
2.1.2. Antagonistic Activity of Bacillus velezensis CE 100 against Pestalotiopsis maculans
2.1.3. Survival Rate of Quercus acutissima Seedlings
2.2. Growth Promotion Effect of Bacillus velezensis CE 100 on Quercus acutissima Seedlings
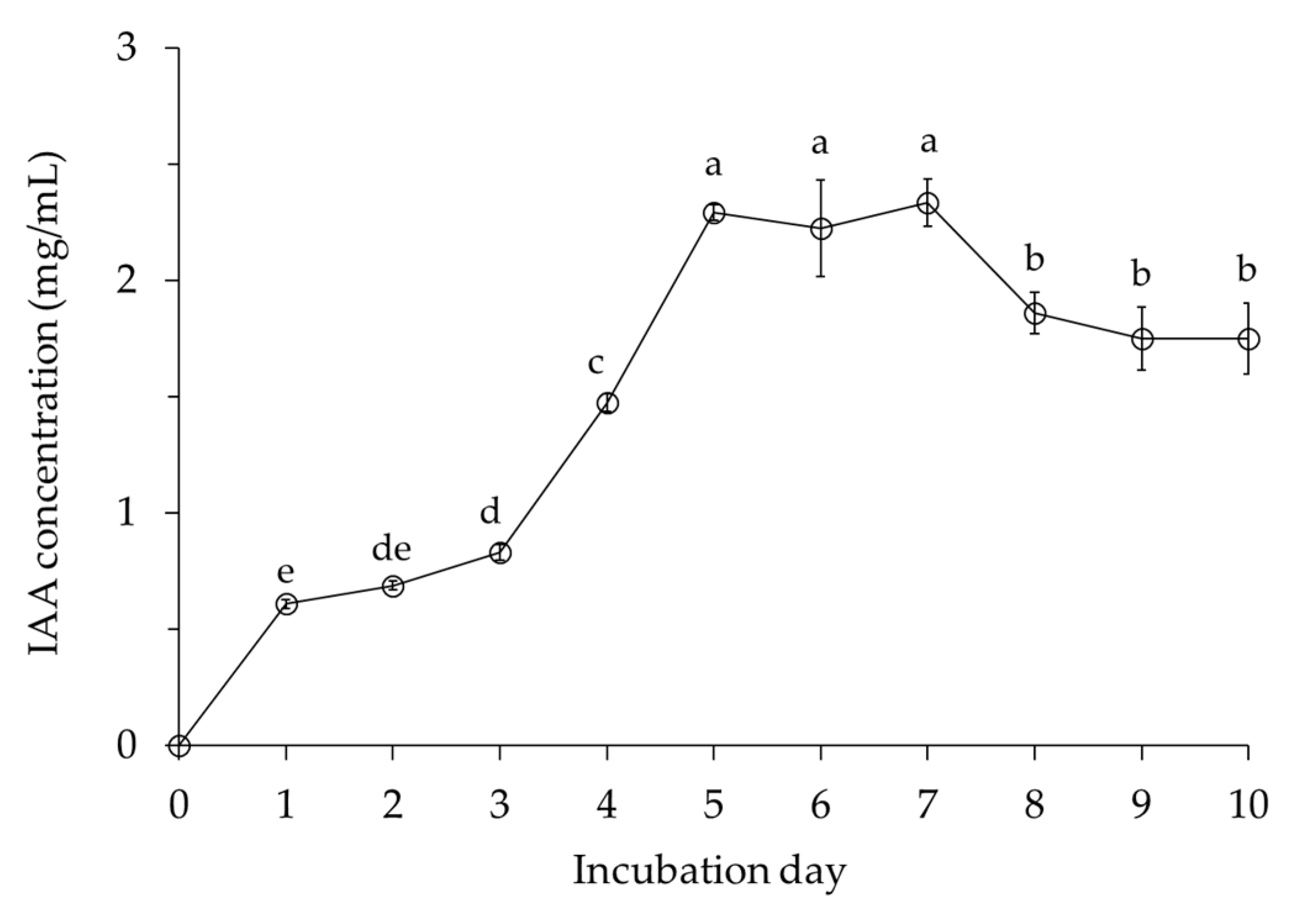
2.2.1. Indole-3-Acetic Acid (IAA) Production
2.2.2. Nutrient Contents in the Growth Media
2.2.3. Nutrient Concentration and Content of Quercus acutissima Seedlings
2.2.4. Biomass of Quercus acutissima Seedlings
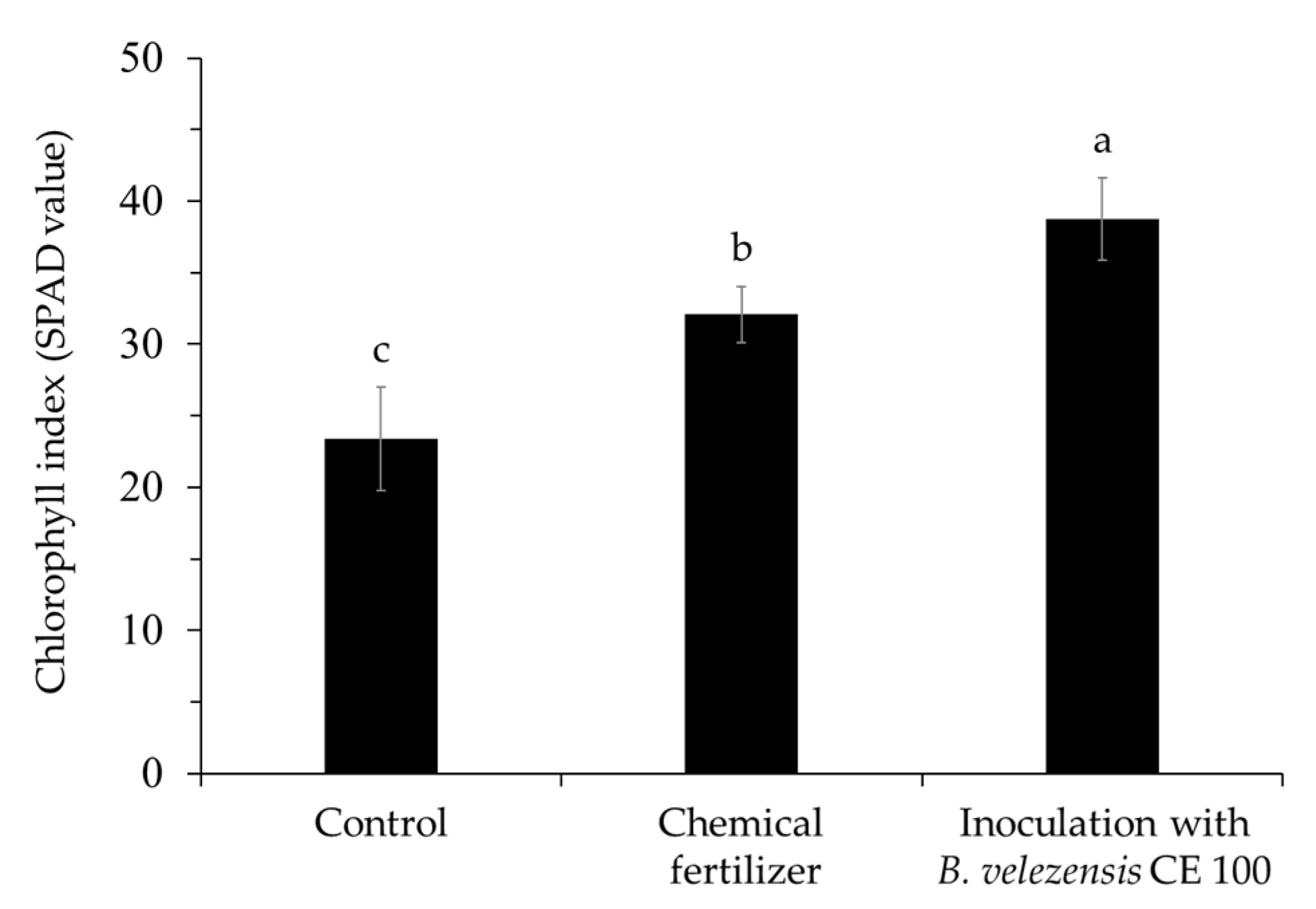
2.2.5. Chlorophyll Index of Quercus acutissima Seedlings
3. Discussion
3.1. Antagonistic Activity of Bacillus velezensis CE 100 against Pestalotiopsis maculans
3.2. Growth Promotion of Quercus acutissima Container Seedlings by Bacillus velezensis CE 100
4. Materials and Methods
4.1. Preparation and Growth Patterns of Bacillus velezensis CE 100
4.2. Isolation and Identification of Pestalotiopsis maculans
4.3. Lytic Enzyme Production by Bacillus velezensis CE 100
4.4. Antagonistic Activity of Bacillus velezensis CE 100 against Pestalotiopsis maculans
4.5. Indole-3-Acetic Acid (IAA) Production by Bacillus velezensis CE 100
4.6. Plant Material and Experimental Conditions
4.7. Growth Medium, Plant Sampling, and Measurement
4.8. Chlorophyll Index of Quercus acutissima Seedlings
4.9. Survival Rate of Quercus acutissima Seedlings
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koenig, W.D.; Mumme, R.L.; Carmen, W.J.; Stanback, M.T. Acorn production by oaks in central coastal California: Variation within and among years. Ecology 1994, 75, 99–109. [Google Scholar] [CrossRef]
- Tanaka, N.; Shimomura, K.; Ishimaru, K. Tannin production in callus cultures of Quercus acutissima. Phytochemistry 1995, 40, 1151–1154. [Google Scholar] [CrossRef]
- Han, Y.; Chang, Y.S. Wood properties and drying characteristics of Korean sawtooth oak (Quercus acutissima Carruth.). Eur. J. Wood Wood Prod. 2020, 78, 1023–1029. [Google Scholar] [CrossRef]
- Chung, Y.J. Comparison of combustion properties of native wood species used for fire pots in Korea. J. Ind. Eng. Chem. 2010, 16, 15–19. [Google Scholar] [CrossRef]
- Youn, Y.-C. The production of oak mushrooms (Lentinula edodes) as a source of farmers income in the Republic of Korea: The case of Cheongyang-Gun. In Forest Products, Livelihoods and Conservation; Center for International Forestry Research: Jakarta, Indonesia, 2004; Volume 1, pp. 94–104. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Eom, M.S.; Yoo, K.S.; Kim, N.C.; Jeon, J.K.; Park, Y.K.; Song, B.H.; Lee, S.H. The yields and composition of bio-oil produced from Quercus acutissima in a bubbling fluidized bed pyrolyzer. J. Anal. Appl. Pyrolysis 2008, 83, 110–114. [Google Scholar] [CrossRef]
- Ryu, S.R.; Choi, H.T.; Lim, J.H.; Lee, I.K.; Ahn, Y.S. Post-fire restoration plan for sustainable forest management in South Korea. Forests 2017, 8, 188. [Google Scholar] [CrossRef] [Green Version]
- Abrams, M.D. Adaptations and responses to drought in Quercus species of North America. Tree Physiol. 1990, 7, 227–238. [Google Scholar] [CrossRef]
- Forestry Statistical Yearbook. Available online: https://www.forest.go.kr/kfsweb/cop/bbs/selectBoardArticle.do;jsessionid=Nz0iyNG4B4kBJtjOhIQQ0yvThcQPtZTF9QxCU93XBxDKK9gYJhZblMP19SFGqLuy.frswas01_servlet_engine5?nttId=3150759&bbsId=BBSMSTR_1064&pageIndex=1&pageUnit=10&searchtitle=title&searchcont=&searchkey=&searchwriter=&searchdept=&searchWrd=&ctgryLrcls=&ctgryMdcls=&ctgrySmcls=&ntcStartDt=&ntcEndDt=&orgId=&mn=NKFS_04_05_09&component= (accessed on 30 June 2021).
- Grossnickle, S.C. Importance of root growth in overcoming planting stress. New Forests 2005, 30, 273–294. [Google Scholar] [CrossRef]
- Tsakaldimi, M.; Zagas, T.; Tsitsoni, T.; Ganatsas, P. Root morphology, stem growth and field performance of seedlings of two Mediterranean evergreen oak species raised in different container types. Plant. Soil 2005, 278, 85–93. [Google Scholar] [CrossRef]
- Wilson, E.R.; Vitals, K.C.; Park, A. Root characteristics and growth potential of container and bare-root seedlings of red oak (Quercus rubra L.) in Ontario, Canada. New Forests 2007, 34, 163–176. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; El-Kassaby, Y.A. Bareroot versus container stocktypes: A performance comparison. New Forests 2016, 47, 1–51. [Google Scholar] [CrossRef]
- Chirino, E.; Vilagrosa, A.; Hernández, E.; Matos, A.; Vallejo, V. Effects of a deep container on morpho-functional characteristics and root colonization in Quercus suber L. seedlings for reforestation in Mediterranean climate. For. Ecol. Manag. 2008, 256, 779–785. [Google Scholar] [CrossRef]
- Juntunen, M.L.; Hammar, T.; Rikala, R. Leaching of nitrogen and phosphorus during production of forest seedlings in containers. J. Environ. Qual. 2002, 31, 1868–1874. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lyu, S.; Sun, D.; Bradshaw, C.J.; Zhou, S. Species decline under nitrogen fertilization increases community-level competence of fungal diseases. Proc. R. Soc. B. 2017, 284, 20162621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilja, A.; Lilja, S.; Kurkela, T. Nursery practices and management of fungal diseases in forest nurseries in Finland: A review. Silva. Fenn. 1997, 31, 79–100. [Google Scholar] [CrossRef] [Green Version]
- Hiddink, G.A.; Termorshuizen, A.J.; van Bruggen, A.H.C. Mixed cropping and suppression of soilborne diseases. In Genetic Engineering, Biofertilisation, Soil Quality and Organic Farming; Springer: Dordrecht, The Netherlands, 2010; Volume 4, pp. 119–146. [Google Scholar] [CrossRef]
- Vettraino, A.; Roques, A.; Yart, A.; Fan, J.T.; Sun, J.H.; Vannini, A. Sentinel trees as a tool to forecast invasions of alien plant pathogens. PLoS ONE 2015, 10, e0120571. [Google Scholar] [CrossRef]
- Yasuda, F.; Kobayashi, T.; Watanabe, H.; Izawa, H. Addition of Pestalotiopsis spp. to leaf spot pathogens of Japanese persimmon. J. Gen. Plant. Pathol. 2003, 69, 29–32. [Google Scholar] [CrossRef]
- Evidente, A.; Zonno, M.C.; Andolfi, A.; Troise, C.; Cimmino, A.; Vurro, M. Phytotoxic α-pyrones produced by Pestalotiopsis guepinii, the causal agent of hazelnut twig blight. J. Antibiot. 2012, 65, 203–206. [Google Scholar] [CrossRef]
- Ren, H.Y.; Li, G.; Qi, X.J.; Fang, L.; Wang, H.R.; Wei, J.G.; Zhong, S. Identification and characterization of Pestalotiopsis spp. causing twig blight disease of bayberry (Myrica rubra Sieb. & Zucc) in China. Eur. J. Plant. Pathol. 2013, 137, 451–461. [Google Scholar] [CrossRef]
- Ray, M.K.; Mishra, P.K.; Baruah, P.K. Control of fungal pathogen Pestalotiopsis disseminata causing grey blight disease in som (Persea bombycina Kost.): An in vitro study. Indian J. Pure Appl. Biosci. 2016, 4, 180–185. [Google Scholar] [CrossRef]
- Monteiro, P.; Gonçalves, M.F.; Pinto, G.; Silva, B.; Martín-García, J.; Diez, J.J.; Alves, A. Three novel species of fungi associated with pine species showing needle blight-like disease symptoms. Eur. J. Plant. Pathol. 2021, 1–20. [Google Scholar]
- Yang, C.; Hamel, C.; Vujanovic, V.; Gan, Y. Fungicide: Modes of action and possible impact on nontarget microorganisms. ISRN Ecol. 2011, 2011, 130289. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.; Sonoda, R. Characterization of moderate resistance to QoI fungicides in Pestalotiopsis longiseta and polymorphism in exon–intron structure of cytochrome b gene. J. Gen. Plant. Pathol. 2012, 78, 398–403. [Google Scholar] [CrossRef]
- Singh, S.; Singh, N.; Kumar, V.; Datta, S.; Wani, A.B.; Singh, D.; Singh, K.; Singh, J. Toxicity, monitoring and biodegradation of the fungicide carbendazim. Environ. Chem. Lett. 2016, 14, 317–329. [Google Scholar] [CrossRef]
- Huang, L.; Li, Q.C.; Hou, Y.; Li, G.Q.; Yang, J.Y.; Li, D.W.; Ye, J.R. Bacillus velezensis strain HYEB5-6 as a potential biocontrol agent against anthracnose on Euonymus japonicus. Biocontrol. Sci. Technol. 2017, 27, 636–653. [Google Scholar] [CrossRef]
- Won, S.-J.; Choub, V.; Kwon, J.-H.; Kim, D.-H.; Ahn, Y.-S. The control of Fusarium root rot and development of coastal pine (Pinus thunbergii Parl.) seedlings in a container nursery by use of Bacillus licheniformis MH48. Forests 2019, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Won, S.-J.; Kwon, J.-H.; Kim, D.-H.; Ahn, Y.-S. The effect of Bacillus licheniformis MH48 on control of foliar fungal diseases and growth promotion of Camellia oleifera seedlings in the coastal reclaimed land of Korea. Pathogens 2019, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, J.-H.; Won, S.-J.; Moon, J.-H.; Kim, C.-W.; Ahn, Y.-S. Control of fungal diseases and increase in yields of a cultivated jujube fruit (Zizyphus jujuba Miller var. inermis Rehder) orchard by employing Lysobacter antibioticus HS124. Forests 2019, 10, 1146. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.-H.; Won, S.-J.; Moon, J.-H.; Lee, U.; Park, Y.-S.; Maung, C.E.H.; Ajuna, H.B.; Ahn, Y.S. Bacillus licheniformis PR2 controls fungal diseases and increases production of jujube fruit under field conditions. Horticulturae 2021, 7, 49. [Google Scholar] [CrossRef]
- Choub, V.; Maung, C.E.H.; Won, S.-J.; Moon, J.-H.; Kim, K.Y.; Han, Y.S.; Cho, J.-Y.; Ahn, Y.S. Antifungal activity of cyclic tetrapeptide from Bacillus velezensis CE 100 against plant pathogen Colletotrichum gloeosporioides. Pathogens 2021, 10, 209. [Google Scholar] [CrossRef]
- Moon, J.-H.; Won, S.-J.; Maung, C.E.H.; Choi, J.-H.; Choi, S.-I.; Ajuna, H.B.; Ahn, Y.S. Bacillus velezensis CE 100 inhibits root rot diseases (Phytophthora spp.) and promotes growth of japanese cypress (Chamaecyparis obtusa Endlicher) seedlings. Microorganisms 2021, 9, 821. [Google Scholar] [CrossRef] [PubMed]
- Free, S.J. Chapter Two—Fungal cell wall organization and biosynthesis. In Advances in Genetics, 1st ed.; Theodore, F., Jay, C.D., Stephen, F.G., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 81, pp. 33–82. [Google Scholar] [CrossRef]
- Shrestha, A.; Sultana, R.; Chae, J.-C.; Kim, K.; Lee, K.-J. Bacillus thuringiensis C25 which is rich in cell wall degrading enzymes efficiently controls lettuce drop caused by Sclerotinia minor. Eur. J. Plant. Pathol. 2015, 142, 577–589. [Google Scholar] [CrossRef]
- Radhapriya, P.; Ramachandran, A.; Palani, P. Indigenous plant growth-promoting bacteria enhance plant growth, biomass, and nutrient uptake in degraded forest plants. 3 Biotech 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Beneduzi, A.; Peres, D.; Vargas, L.K.; Bodanese-Zanettini, M.H.; Passaglia, L.M.P. Evaluation of genetic diversity and plant growth promoting activities of nitrogen-fixing Bacilli isolated from rice fields in South Brazil. Appl. Soil Ecol. 2008, 39, 311–320. [Google Scholar] [CrossRef]
- Ahmed, A.; Hasnain, S. Auxins as one of the factors of plant growth improvement by plant growth promoting rhizobacteria. Pol. J. Microbiol. 2014, 63, 261–266. [Google Scholar] [CrossRef]
- Mehta, P.; Walia, A.; Kulshrestha, S.; Chauhan, A.; Shirkot, C.K. Efficiency of plant growth-promoting P-solubilizing Bacillus circulans CB7 for enhancement of tomato growth under net house conditions. J. Basic Microbiol. 2015, 55, 33–44. [Google Scholar] [CrossRef]
- Enders, T.A.; Strader, L.C. Auxin activity: Past, presentand future. Am. J. Bot. 2015, 102, 180–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuan, K.B.; Othman, R.; Abdul Rahim, K.; Shamsuddin, Z.H. Plant growth-promoting rhizobacteria inoculation to enhance vegetative growth, nitrogen fixation and nitrogen remobilisation of maize under greenhouse conditions. PLoS ONE 2016, 11, e0152478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Newman, E.I.; Andrews, R.E. Uptake of phosphorus and potassium in relation to root growth and root density. Plant. Soil 1973, 38, 49–69. [Google Scholar] [CrossRef]
- Salifu, K.F.; Apostol, K.G.; Jacobs, D.F.; Islam, M.A. Growth, physiology, and nutrient retranslocation in nitrogen-15 fertilized Quercus rubra seedlings. Ann. For. Sci. 2008, 65, 101. [Google Scholar] [CrossRef] [Green Version]
- Nicodemus, M.A.; Salifu, F.K.; Jacobs, D.F. Growth, nutrition, and photosynthetic response of black walnut to varying nitrogen sources and rates. J. Plant. Nutr. 2008, 31, 1917–1936. [Google Scholar] [CrossRef]
- Bassi, D.; Menossi, M.; Mattiello, L. Nitrogen supply influences photosynthesis establishment along the sugarcane leaf. Sci. Rep. 2018, 8, 2327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, J.H.; Kim, S.-H.; Khaine, I.; Kwak, M.J.; Lee, H.K.; Lee, T.Y.; Lee, W.Y.; Woo, S.Y. Physiological changes and growth promotion induced in poplar seedlings by the plant growth-promoting rhizobacteria Bacillus subtilis JS. Photosynthetica 2018, 56, 1188–1203. [Google Scholar] [CrossRef]
- Łakomy, P.; Kuźmiński, R.; Mucha, J.; Zadworny, M. Effects of oak root pruning in forest nurseries on potential pathogen infections. For. Pathol. 2019, 49, e12513. [Google Scholar] [CrossRef]
- Dahiya, N.; Tewari, R.; Hoondal, G.S. Biotechnological aspects of chitinolytic enzymes: A review. Appl. Microbiol. Biotechnol. 2006, 71, 773–782. [Google Scholar] [CrossRef]
- Geoghegan, I.; Steinberg, G.; Gurr, S. The role of the fungal cell wall in the infection of plants. Trends Microbiol. 2017, 25, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Choub, V.; Ajuna, H.B.; Won, S.-J.; Moon, J.-H.; Choi, S.-I.; Maung, C.E.H.; Kim, C.-W.; Ahn, Y.S. Antifungal activity of Bacillus velezensis CE 100 against anthracnose disease (Colletotrichum gloeosporioides) and growth promotion of walnut (Juglans regia L.) trees. Int. J. Mol. Sci. 2021, 22, 10438. [Google Scholar] [CrossRef]
- Uchida, R. Essential Nutrients for Plant Growth: Nutrient Functions and Deficiency Symptoms. In Plant Nutrient Management in Hawaii’s Soils; Silva, J.A., Uchida, R., Eds.; College of Tropical Agriculture and Human Resources, University of Hawaii at Manoa: Hawaii, HI, USA, 2000; Volume 4, pp. 31–55. [Google Scholar]
- Choi, T.G.; Maung, C.E.H.; Lee, D.R.; Henry, A.B.; Lee, Y.S.; Kim, K.Y. Role of bacterial antagonists of fungal pathogens, Bacillus thuringiensis KYC and Bacillus velezensis CE 100 i n control of root-knot nematode, Meloidogyne incognita and subsequent growth promotion of tomato. Biocontrol. Sci. Technol. 2020, 30, 685–700. [Google Scholar] [CrossRef]
- Lingappa, Y. Chitin media for selective isolation and culture of Actinomycetes. Phytopathology 1962, 52, 317–323. [Google Scholar]
- Skidmore, A.; Dickinson, C. Colony interactions and hyphal interference between septoria nodorum and phylloplane fungi. Trans. Brit. Mycol. Soc. 1976, 66, 57–64. [Google Scholar] [CrossRef]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bremner, J.M. Determination of nitrogen in soil by the Kjeldahl method. J. Agric. Sci. 1960, 55, 11–33. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Growth Media | Seedling | ||||
|---|---|---|---|---|---|---|
| Nutrient Content (g/kg) | Nutrient Concentration (%/plant) | Nutrient Content (mg/plant) | ||||
| Total Nitrogen | Total Phosphorus | Total Nitrogen | Total Phosphorus | Total Nitrogen | Total Phosphorus | |
| Control | 2.63 ± 0.14 b | 0.19 ± 0.01 c | 0.39 ± 0.08 c | 0.10 ± 0.01 c | 303.47 ± 45.98 c | 79.00 ± 1.02 c |
| Chemical fertilizer | 2.98 ± 0.08 a | 0.57 ± 0.09 a | 0.59 ± 0.06 b | 0.14 ± 0.02 b | 722.35 ± 117.59 b | 147.66 ± 1.01 b |
| Bacterial inoculation | 2.85 ± 0.16 a | 0.44 ± 0.05 b | 0.74 ± 0.03 a | 0.22 ± 0.04 a | 931.71 ± 54.96 a | 370.10 ± 2.76 a |
| Treatment | Shoot Dry Mass (g/plant) | Root Dry Mass (g/plant) | Total Dry Mass (g/plant) |
|---|---|---|---|
| Control | 35.76 ± 6.45 c | 43.85 ± 7.63 c | 76.62 ± 13.69 c |
| Chemical fertilizer | 68.09 ± 6.95 b | 54.52 ± 12.05 b | 122.61 ± 13.67 b |
| Bacterial inoculation | 88.22 ± 8.79 a | 76.42 ± 13.57 a | 164.64 ± 16.24 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Won, S.-J.; Moon, J.-H.; Ajuna, H.B.; Choi, S.-I.; Maung, C.E.H.; Lee, S.; Ahn, Y.S. Biological Control of Leaf Blight Disease Caused by Pestalotiopsis maculans and Growth Promotion of Quercus acutissima Carruth Container Seedlings Using Bacillus velezensis CE 100. Int. J. Mol. Sci. 2021, 22, 11296. https://doi.org/10.3390/ijms222011296
Won S-J, Moon J-H, Ajuna HB, Choi S-I, Maung CEH, Lee S, Ahn YS. Biological Control of Leaf Blight Disease Caused by Pestalotiopsis maculans and Growth Promotion of Quercus acutissima Carruth Container Seedlings Using Bacillus velezensis CE 100. International Journal of Molecular Sciences. 2021; 22(20):11296. https://doi.org/10.3390/ijms222011296
Chicago/Turabian StyleWon, Sang-Jae, Jae-Hyun Moon, Henry B. Ajuna, Su-In Choi, Chaw Ei Htwe Maung, Sangtae Lee, and Young Sang Ahn. 2021. "Biological Control of Leaf Blight Disease Caused by Pestalotiopsis maculans and Growth Promotion of Quercus acutissima Carruth Container Seedlings Using Bacillus velezensis CE 100" International Journal of Molecular Sciences 22, no. 20: 11296. https://doi.org/10.3390/ijms222011296